Findings
Motor & Sensory nerve responses
Motor Responses
The motor response is obtained by stimulating a nerve and recording from a muscle that it innervates. The muscle selected should have a fairly well-defined motor point, and preferably be relatively isolated from other muscles innervated by the nerve and from other nerves that may be stimulated inadvertently during the test. The excitation of nearby muscles may alter the response and make it difficult to determine the exact onset of the desired motor response.
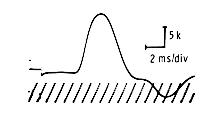
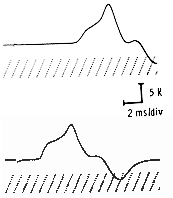
The motor response may be characterized by its amplitude, duration, and wave form. The amplitude is measured from the baseline to the top of the negative peak of the motor response and is expressed in millivolts.
The distal latency is measured from the onset of the stimulus artifact to the point of takeoff from the baseline and is measured in milliseconds. Extra care must be taken to use the corresponding takeoff points of both the distal and proximal responses so that conduction velocities are measured along the same fibers. The amplitude depends to a large extent on the number and size of muscle fibers being activated, and supramaximal stimulation of the nerve should ensure a maximal motor response. Any pathological process that decreases the number of motor units or muscle fibers responding will affect the amplitude. The normal motor response indicates a fairly synchronous discharge of the motor units. If there is dispersion of the times when the motor units discharge, then the amplitude will be lowered and the response spread in time. This effect brings up the question of duration of the response. In processes in which the nerve conduction slows differentially, the duration of the response will be prolonged and thus its amplitude decreased.
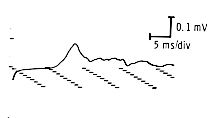
The usual motor response has a fairly simple waveform. It may have one or two initial negative (up) peaks (the latter usually indicating two muscle being stimulated) and usually will be followed by a positive deflection (down) toward the end. The response should have a clear initial negative deflection as it takes off from the baseline. In some pathological processes, the wave may have multiple phases, appearing extremely complex.
The motor response also changes in relationship to the point of nerve stimulation. The more proximally the nerve is stimulated, the lower the amplitude and the longer the duration of responses seen. These effects are due to the temporal dispersion of the motor units activated because of differential conduction velocities in the normal motor nerves.
Sensory-Nerve Action Potentials
Sensory-nerve action potentials (NAP) are obtained by stimulating a nerve and recording directly from it or one of its branches. The recording site must be remote from muscles innervated by that same nerve because muscle responses will obscure the much smaller NAP.
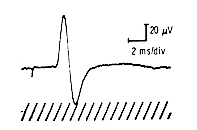
The NAP can also be characterized by it amplitude, duration, and wave form. The amplitude of the NAP is measured from the peak of the positive deflection the peak of the negative deflection and is measured in microvolts. The sensory distal latency is traditionally measured from the stimulus artifact to the takeoff or the peak of the negative deflection. When conduction velocities are needed, distal latencies to the takeoff of the proximal and distal responses should be used. The amplitude depends on the number of axons being stimulated and the synchrony with which they transmit their impulse. If the axons transmit impulses at comparable velocities, the response duration will be short and amplitude high. However, if the axonal velocities are widely dispersed, the NAP duration will be longer and its amplitude lower.
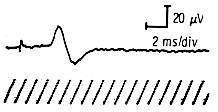
Distal Latency
Defined as the time from the stimulus affecting the nerve to the response (motor or sensory) being recorded, latency is usually measured in milliseconds (msec). Distal latency is that interval measured from the stimulation of the distal-most accessible site on the nerve. This finding does not give direct information on conduction velocities, because the distal segment often follows a tortuous route that cannot be measured. The measurement is useful, however, because it can be compared with normal data and indicate the relative conductivity of the segment of the nerve. In measuring the latency of the motor nerve, remember that a small portion of that time is due to the delay in neuromuscular transmission, whereas no such delay is present in sensory latencies.
Conduction Velocity
If a nerve can be stimulated at two points along its course, and a measurement can be obtained of the distance between those points, conduction velocities can be figured.
This is true for most motor nerves. In sensory studies however, only one stimulation site is nromally used. Compute the velocity (V) by measuring the distance (d) in millimeters (mm) between the two stimulation points and dividing by the difference in latency (ms) between the proximal (tp) and distal stimulation points (td), as indicated in this equation:
V=d/tp-td
The result is expressed as meters per second (m/sec.).
Because the proximal and distal latencies are measured to the takeoff of the response, the conduction velocity obtained represents conductions along the fastest conducting fibers, with those that first reach the muscle causing the initial deflection.
Conduction velocities in the various nerves differ, depending on anatomical considerations. However, several general principles apply to evaluating nerve conduction studies:
The more proximal the segment of nerve being evaluated is, the faster the velocity will be. If the extremity being tested is cold, the velocity will be slowed and the amplitude increased. This effect occurs especially in cold weather and some provisions for warming the patient and for using a fairly constant room temperature should be made. At times anatomical considerations such as potential entrapment points will also tend to slow the velocities. The shorter the segment between the two stimulation points, the less reliable the calculated velocities will be, due to a greater effect on the margin of error by a shorter distance.
Conduction velocities depend most on the integrity of the myelin sheath. In segmental demyelinating diseases conduction velocities drop to below 50 percent of normal values. However, when axonal loss is severe, the velocity will also be slowed due to a dropout of the fastest conducting fibers. The drop in axonal loss is usually in the vicinity of 30 percent below normal values.
Machine Settings
In the study of sensory and motor responses, different filter, sweep speed, and sensitivity settings are used. Sensory studies are performed with the low frequency setting between 32 and 50 Hz and the high frequencies between 1.6-2 and 3 KHz. The sweep speed is set to 2 ms/division and the sensitivity at 10-20 µV/division. Motor studies are performed with the low frequency set to 1.6-2 Hz and the high frequencies to 8-10 KHz. Depending on the response’s latency and duration, the sweep speed can be set to anywhere between 2-5 ms/division and the sensitivity between 2-10 mv/division. Whatever the setting, the distal and proximal latencies should be measured at the same setting, preferably using the faster sweep speed, as the takeoff is easier to identify with faster sweeps.
Normal Values
Normal values can be sorted according to age, sex, extremity length, patient’s height or a combination thereof. Unless otherwise specified, we use the Cleveland Clinic Foundation’s EMG Lab normal values which were sorted according to patient’s age. These normals were based on a sampling of a minimum of forty patients for ages ten to nineteen, and seventy and over, and at least ninety patients for the other age-groups. The ranges (first two numbers) and averages (between parenthesis) are provided. These values are based on the following standard distances: 13 cm for the median sensory (wrist to active electrode), 11 cm for the ulnar sensory and 10 cm for the radial sensory. For the motor studies, a minimum of 4-6 cm is used between the wrist and active electrodes (median and ulnar nerves).